Towards a Programmable Material
Radhika Nagpal
Amorphous
Computing Group
MIT Artificial Intelligence Lab
(1999, Slide Show)
Imagine a flexible substrate, consisting of millions of tiny
interwoven programmable fibers, that can be programmed to assume a
large variety of global shapes. Rather than build precisely engineered
mechanical structures, a programmable material would allow one to
program precise complicated structures starting from a single flexible
mechanical base. Not only could one design many complex static
structures by programming a single substrate, but also create dynamic
structures that react to, and affect, the environment. For example, a
flexible car surface that can change structure exactly at the point of
impact, rather than having specifically engineered crumple points; an
airplane wing that can dynamically change shape to resist shear; a
programmable assembly line that can move objects around by producing
ripples in specific directions; a reconfigurable robot that changes it
shape based on what function it needs to perform; or a manufacturing
line that replaces precise mechanical engineering with programming.
Programmable materials would make possible a host of novel
applications that blur the boundary between computation and the
environment.
Biology, in particular morphogenesis, gives us many examples of
complex structures formed from the cooperation of large numbers of
cells. One example are epithelial cell sheets that can organize
into complex structures such as branching tubes, neural folds and
other structures through the actuation of many individual epithelial
cells, each cell having limited force. Here we propose a model
inspired by epithelial cell sheets - the programmable material is
composed of identically programmed, connected, mechanical cells that
organize into complex structures through the coordination of local
shapes changes.
A Single Active Cell
Odell, Oster, Alberch, and Burnside[1] present a mechanical model for how
embryos form structures during neurulation and gastrulation, through the
local deformations of epithelial cells. Each cell consists of fibers and
contains a slightly compressible fluid. Apical (top) fibers can actively
contract, and cause what is known as a purse string contraction.
The cells are connected to each other and can cause invaginations when
several neighboring cells contract.
We use the cell model proposed by Odell et al, but allow all fibers
to actively contract, or loosen. By programming the active fibers, one
can create many simple shape changes - long columnar cells, short fat cells,
triangular or conical cells. When these cells are connected together, many
global
shapes can be expressed, simply as
compositions
of these local shapes. |
| Various single cell deformations |
  |
  |
|
Structures Composed of Active Cells
The active cells can be connected together to form a flexible
substrate which can then be programmed to form different global
structures.
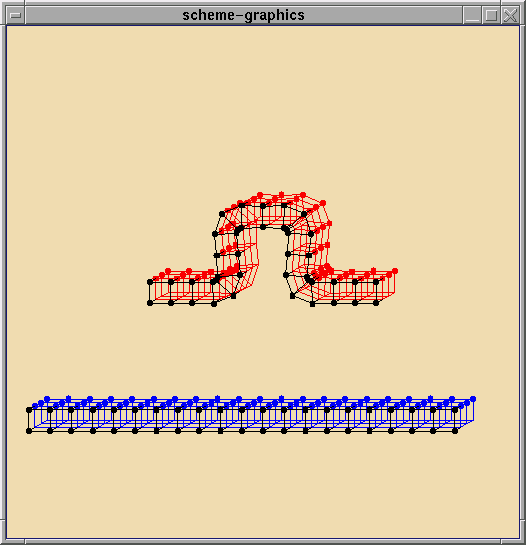
Structure I: A Ring of Active Cells
Here we present some biologically inspired shapes formed by a ring of cells.
The 2D ring can represent a sphere or cylinder. These are not meant to
model exact biology. In fact practically any shape can be formed, given
the limitation of the number of cells and that they are connected in a
ring. Understanding the underlying general organizational principles for
reliable global behavior present in biological systems will have important
implications for applications that embed computation into the environment.
Each of these shapes can be described as simple local shape changes
by individual cells. Combining this with simple
pattern formation techniques (chemical tropisms, coordinate systems,etc)
from amorphous computing, one can program
the cells to coordinate to form the desired mechanical pattern.
| Gastrulation and Ventral Furrow Formation:
The first example approximates gastrulation in organisms such as a sea
urchin. The active cells contract their apical and lateral fibers, with
more contraction happening at the tip of the invagination. Eventually the
invagination goes all the way through the body to form the gut. The second
picture shows ventral furrow formation in drosophilia, where cells use
apical contraction to form a tube on the ventral side of the cylinderical
embryo (similar to example in [1]). |
 |
 |
| Cleaving into Two and the formation of Eyefolds: In a manner
similar to gastrulation, the ring (or sphere) can form two smaller rings.
This could represent the cleaving of a cell through the actions of fibers
distributed in the cell membrane. The second example shows the formation
of folds in the ring through alternating cells with apical contractions
and basal contractions. Folds are common during development (chick cornea),
however usually due to differential growth. |
 |
 |
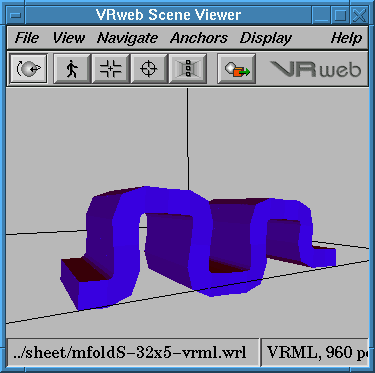
| Bulb Formation, and Three fingers: Bulb formation is simply
the opposite of ventral furrow. The fingers are also similar to bulbs,
however with deepening invaginations (similar mechanisms to gastrulation)
between the fingers. |
 |
 |
Programming the Ring of Cells
Several languages have been developed for programming amorphous algorithms
at the level of the individual processors. For this example we use a language
developed by Weiss for programming biological cells. The main advantage
of this language is it provides a very simple way for expressing the program
for a single cell and incorporates many amorphous computing ideas.
The program for a cell consists of a unordered set of independent rules
and each rule consists consists of a triggering event, a condition and
actions. The state of a cell consists of boolean markers and timers.
Events consist of recieving a message from a neighboring cell, a marker
whose timer expired or a sensor triggering. The condition is a boolean
expression of markers. The possible actions are to affect the state, send
a message, and set actuators. Underlying this language are gradients (see
self-organising coordinate systems). that can be used to mark regions.
All inter-cellular communication is through gradients.
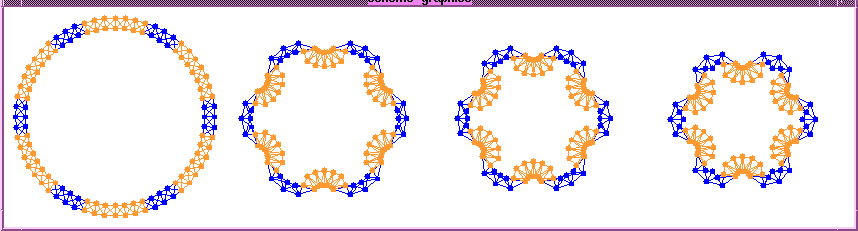
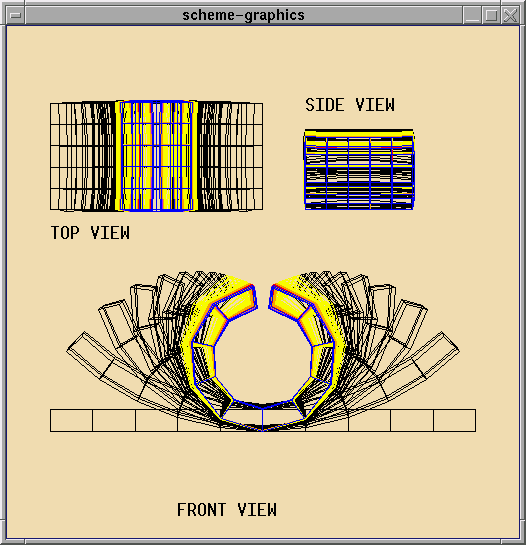
A Cell Program for Initiating Eyefolds:
The result of this program is that folds (or invaginations) appear
at random places on the ring with a separation of between 3 to 6 cells.
The program uses the clubs algorithm (see
publications), which uses short range inhibition
to ensure that folds appear with some preferred spacing.
The program can be extended to cause cells close to the fold points
to also contract their apical fibers. This program generates folds that
are slightly different from this simulation example, where the folds are
equidistant.
The language currently does not have all the features necessary to implement
general shapes. Other amorphous ideas, such as tropism, gradient slopes,
etc can be added to allow one to express cell programs for all of the shapes
mentioned above. |

 |
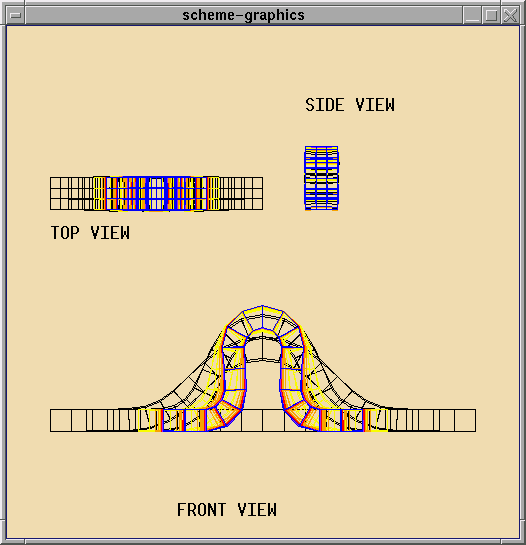
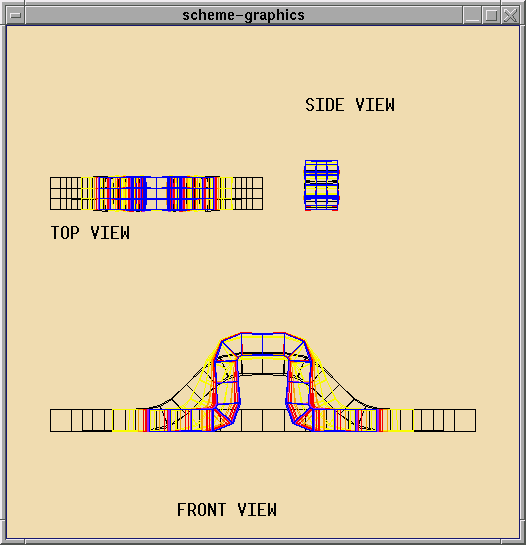
Structure II: A Sheet of Active Cells

The shapes that can be formed with a ring are limited. A more interesting
structure is a flexible sheet. One can imagine a wide
variety of shapes that can achieved by folding the sheet in various
ways. Currently the focus is on experimenting with different basic
shapes and studying limitations on shapes that can be achieved.
The next step is to determine mechanisms for composing
basic shapes to produce more complex static
and dynamic shapes. For example folds could be composed to generate
more complex geometric structures - cubes, ridges, or, more ambitiously
origami. Another area of investigation is dynamic shapes like a ring that
locomotes by extending pseudopodia or traveling waves on the cell sheet
that result either in movement of the sheet or movement of an object on
the sheet.
This project is part of the
amorphous
computing project[2]. The challenge is to identify general principles
for achieving coherent and reliable global behavior from the local interactions
of large numbers of programmable parts. Each part has limited resources,
limited reliablity and only local sensory information and limited-range
actuator impact. The goal of the amorphous computing project is both to
identify new parallel programming paradigms and to invent the computational
substrates that can support this kind of engineering.
References
[1] The Mechanical Basis of Morphogenesis, 1. Epithelial Folding and
Invagination, G.M. Odell, G. Oster, P.Alberch, B. Burnside, Developmental
Biology, vol 85, 1981.
[2] Amorphous Computing, H. Abelson, T. Knight and G. Sussman,
white-paper Oct 1995.